Spawn is the eggs and sperm released or deposited into water by aquatic animals. As a verb, to spawn refers to the process of freely releasing eggs and sperm into a body of water ; the physical act is known as spawning. The vast majority of aquatic and amphibious animals reproduce through spawning. These include the following groups:Bony and cartilaginous fishes
Crustaceans
Mollusks
Echinoderms
Amphibians
Aquatic insects
Coral, which are living colonies of tiny, aquatic organisms—not plants, as they are sometimes perceived to be. Corals, while appearing sedentary or botanical by nature, actually spawn by releasing clouds of sperm and egg cells into the water column, where the two mix.
The spawn (eggs) of a clownfish. The black spots are the developing eyes.
Pacific salmon are semelparous or "big bang" spawners, which means they die shortly after spawning
The pickled and dehydrated roe of mullet
Cutthroat trout are monogamous pair spawners
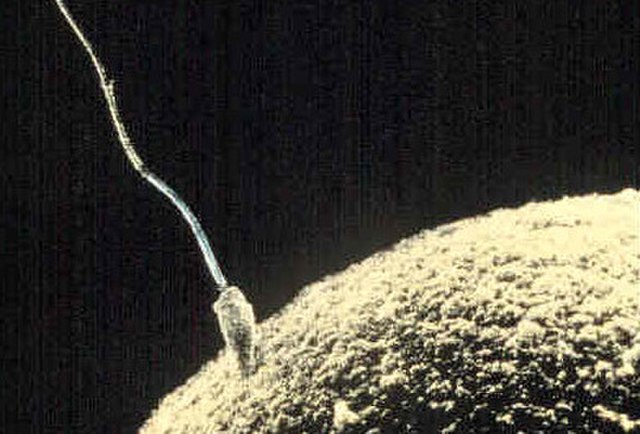
The egg cell or ovum is the female reproductive cell, or gamete, in most anisogamous organisms. The term is used when the female gamete is not capable of movement (non-motile). If the male gamete (sperm) is capable of movement, the type of sexual reproduction is also classified as oogamous. A nonmotile female gamete formed in the oogonium of some algae, fungi, oomycetes, or bryophytes is an oosphere. When fertilized, the oosphere becomes the oospore.
Ovum and sperm fusing together
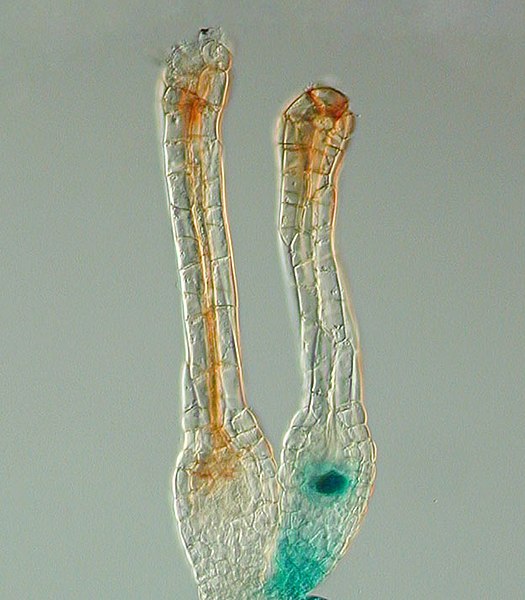
Gene expression pattern determined by histochemical GUS assays in Physcomitrella patens. The Polycomb gene FIE is expressed (blue) in unfertilized egg cells of the moss Physcomitrella patens (right) and expression ceases after fertilization in the developing diploid sporophyte (left). In situ GUS staining of two female sex organs (archegonia) of a transgenic plant expressing a translational fusion of FIE-uidA under control of the native FIE promoter