Afroinsectiphilia
Videos
Page
The Afroinsectiphilia is a clade that has been proposed based on the results of recent molecular phylogenetic studies. Many of the taxa within it were once regarded as part of the order Insectivora, but Insectivora is now considered to be polyphyletic and obsolete. This proposed classification is based on molecular studies only, and there is no morphological evidence for it.

Afroinsectiphilia
Potamogalidae
Videos
Page
Potamogalidae is the family of "otter shrews", a group of semiaquatic riverine afrotherian mammals indigenous to sub-Saharan Africa. They are most closely related to the tenrecs of Madagascar, from which they are thought to have split about 47–53 million years ago. They were formerly considered a subfamily of Tenrecidae.

Image: Potamogale chaillu
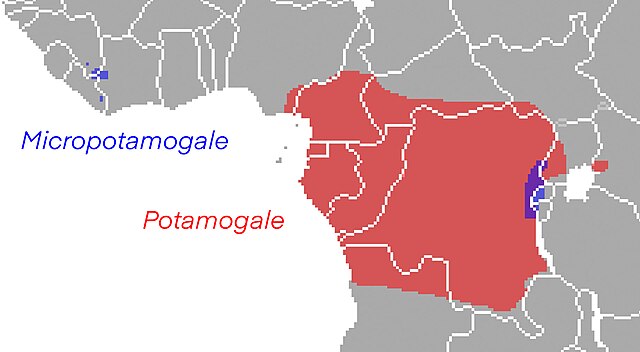
Image: Potamogalidae range