Archaea
Videos
Archaea is a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotic. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.

Archaea were discovered in volcanic hot springs. Pictured here is Grand Prismatic Spring of Yellowstone National Park.

The ARMAN are a group of archaea discovered in acid mine drainage in the early 2000s.
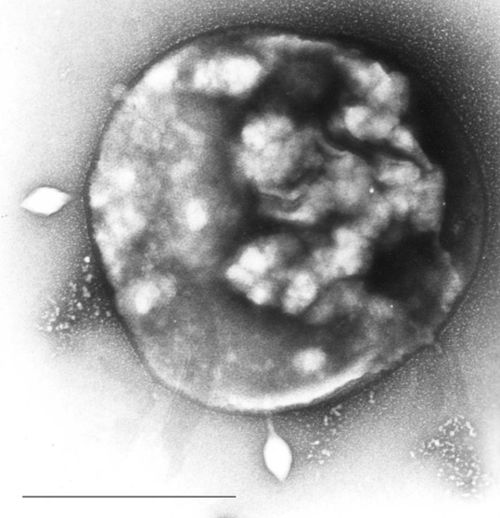
Sulfolobus infected with the DNA virus STSV1. Bar is 1 micrometer.

Archaea that grow in the hot water of the Morning Glory Hot Spring in Yellowstone National Park produce a bright colour
Haloquadratum walsbyi
Videos
Haloquadratum walsbyi is a species of Archaea in the genus Haloquadratum, known for its square shape and halophilic nature.


Microscopic image from the hypersaline Lake Tyrrell, in which orange chlorophyte Dunaliella salina can be tentatively identified, accompanied by a number of smaller Haloquadratum walsbyi, showing their flat square-shaped cells.