Bacillus anthracis
Videos
Bacillus anthracis is a gram-positive and rod-shaped bacterium that causes anthrax, a deadly disease to livestock and, occasionally, to humans. It is the only permanent (obligate) pathogen within the genus Bacillus. Its infection is a type of zoonosis, as it is transmitted from animals to humans. It was discovered by a German physician Robert Koch in 1876, and became the first bacterium to be experimentally shown as a pathogen. The discovery was also the first scientific evidence for the germ theory of diseases.

CD14 and Mac-1 work together to increase the macrophage internalization of B. anthracis spores. (a) CD14 and TLR2 work in close proximity to propagate the activation signal. Mac-1 starts in a closed confirmation which keeps the integrin from binding the spores, keeping them out of the cell. (b) CD14 binds the rhamnose residues on the BclA of the exosporium leading to TLR2-mediated activation of PI3K. (c) Activation of PI3K pushes Mac-1 into its open conformation so the spore can be bound and internalized (information from Oliva

CapD protein crystal structure of B. anthracis
Neutrophil
Videos
Neutrophils are a type of white blood cell. More specifically, they form the most abundant type of granulocytes and make up 40% to 70% of all white blood cells in humans. They form an essential part of the innate immune system, with their functions varying in different animals.
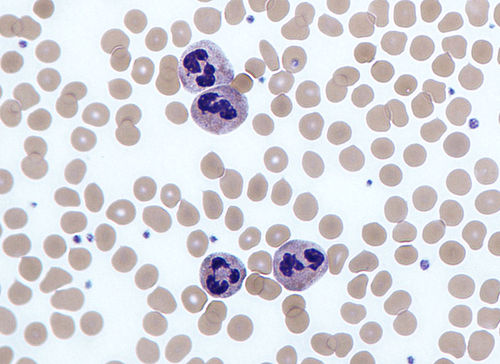
Neutrophils with segmented nuclei surrounded by erythrocytes and platelets. Intra-cellular granules are visible in the cytoplasm (Giemsa stained).

Hypersegmented neutrophil

Scanning electron micrograph of a neutrophil (yellow) phagocytosing anthrax bacilli (orange). Scale bar is 5 μm.

Micrograph showing several neutrophils during an acute inflammation