Belemnitida is an extinct order of squid-like cephalopods that existed from the Late Triassic to Late Cretaceous. Unlike squid, belemnites had an internal skeleton that made up the cone. The parts are, from the arms-most to the tip: the tongue-shaped pro-ostracum, the conical phragmocone, and the pointy guard. The calcitic guard is the most common belemnite remain. Belemnites, in life, are thought to have had 10 hooked arms and a pair of fins on the guard. The chitinous hooks were usually no bigger than 5 mm (0.20 in), though a belemnite could have had between 100 and 800 hooks in total, using them to stab and hold onto prey.
Belemnitida
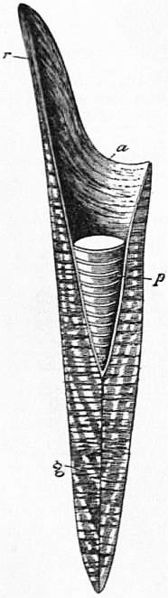
Cone diagram: r pro-ostracum, a alveolus, p phragmocone, g guard
Reconstruction of a typical belemnite
The Jurassic Youngibelus reconstructed with hectocotyli
A cephalopod is any member of the molluscan class Cephalopoda such as a squid, octopus, cuttlefish, or nautilus. These exclusively marine animals are characterized by bilateral body symmetry, a prominent head, and a set of arms or tentacles modified from the primitive molluscan foot. Fishers sometimes call cephalopods "inkfish", referring to their common ability to squirt ink. The study of cephalopods is a branch of malacology known as teuthology.
Cephalopod
The primitive nautilus eye functions similarly to a pinhole camera.
A cuttlefish with W-shaped pupils which may help them discern colors
This broadclub cuttlefish (Sepia latimanus) can change from camouflage tans and browns (top) to yellow with dark highlights (bottom) in less than a second.