Brain
Videos
The brain is an organ that serves as the center of the nervous system in all vertebrate and most invertebrate animals. In vertebrates, a small part of the brain called the hypothalamus is the neural control center for all endocrine systems. The brain is the largest cluster of neurons in the body and is typically located in the head, usually near organs for special senses such as vision, hearing and olfaction.
It is the most energy-consuming organ of the body, and the most specialized, responsible for endocrine regulation, sensory perception, motor control, and the development of intelligence.


Cross section of the olfactory bulb of a rat, stained in two different ways at the same time: one stain shows neuron cell bodies, the other shows receptors for the neurotransmitter GABA.
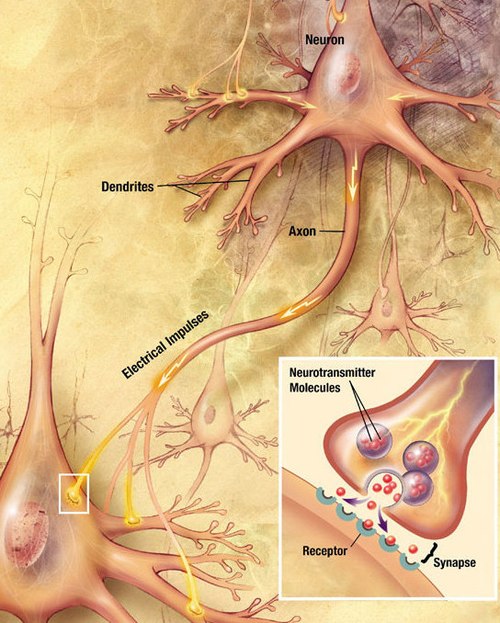
Neurons generate electrical signals that travel along their axons. When a pulse of electricity reaches a junction called a synapse, it causes a neurotransmitter chemical to be released, which binds to receptors on other cells and thereby alters their electrical activity.

Fruit flies (Drosophila) have been extensively studied to gain insight into the role of genes in brain development.
Head
Videos
A head is the part of an organism which usually includes the ears, brain, forehead, cheeks, chin, eyes, nose, and mouth, each of which aid in various sensory functions such as sight, hearing, smell, and taste. Some very simple animals may not have a head, but many bilaterally symmetric forms do, regardless of size.

The head of a meerkat

The head of a death mask

Mid-sagittal section of a human skull, by Leonardo da Vinci, c. 1489

Transection of a human head, by Thomas Bartholin, 1673