Brown algae
Videos
Photos
Brown algae are a large group of multicellular algae comprising the class Phaeophyceae. They include many seaweeds located in colder waters of the Northern Hemisphere. Brown algae are the major seaweeds of the temperate and polar regions. Many brown algae, such as members of the order Fucales, commonly grow along rocky seashores. Most brown algae live in marine environments, where they play an important role both as food and as a potential habitat. For instance, Macrocystis, a kelp of the order Laminariales, may reach 60 m (200 ft) in length and forms prominent underwater kelp forests that contain a high level of biodiversity. Another example is Sargassum, which creates unique floating mats of seaweed in the tropical waters of the Sargasso Sea that serve as the habitats for many species. Some members of the class, such as kelps, are used by humans as food.

Brown algae

Two specimens of Laminaria hyperborea, each showing the rootlike holdfast at lower left, a divided blade at upper right, and a stemlike stipe connecting the blade to the holdfast.

Species like Fucus vesiculosus produce numerous gas-filled pneumatocysts (air bladders) to increase buoyancy.

Growth in Dictyota dichotoma occurs at each frond tip, where new cells are produced.
Multicellular organism
Videos
Photos
A multicellular organism is an organism that consists of more than one cell, in contrast to unicellular organism. All species of animals, land plants and most fungi are multicellular, as are many algae, whereas a few organisms are partially uni- and partially multicellular, like slime molds and social amoebae such as the genus Dictyostelium.
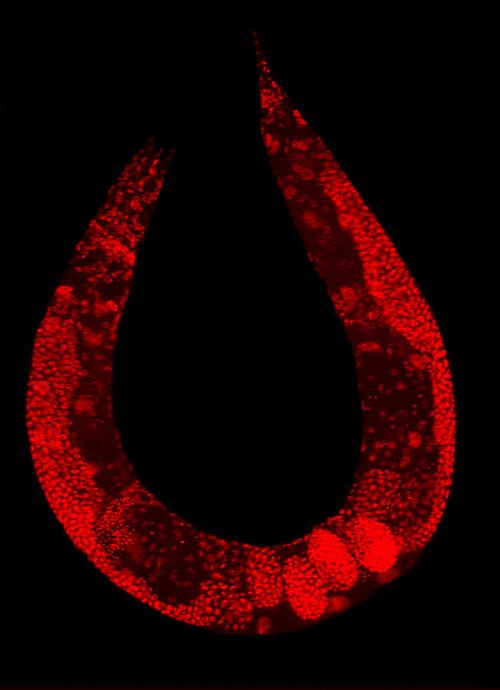
The nematode Caenorhabditis elegans stained to highlight the nuclei of its cells