Cell nucleus
Videos
The cell nucleus is a membrane-bound organelle found in eukaryotic cells. Eukaryotic cells usually have a single nucleus, but a few cell types, such as mammalian red blood cells, have no nuclei, and a few others including osteoclasts have many. The main structures making up the nucleus are the nuclear envelope, a double membrane that encloses the entire organelle and isolates its contents from the cellular cytoplasm; and the nuclear matrix, a network within the nucleus that adds mechanical support.
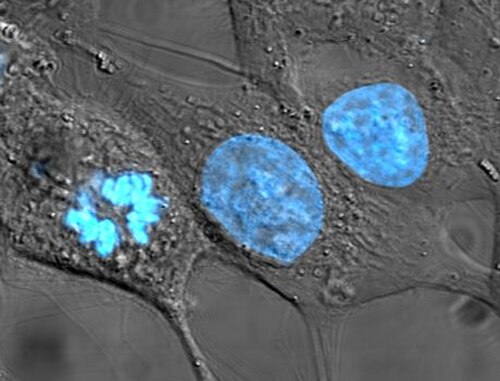
HeLa cells stained for nuclear DNA with the blue fluorescent Hoechst dye. The central and rightmost cells are in interphase, thus their entire nuclei are labeled. On the left, a cell is going through mitosis and its DNA has condensed.

An image of a newt lung cell stained with fluorescent dyes during metaphase. The mitotic spindle can be seen, stained green, attached to the two sets of chromosomes, stained light blue. All chromosomes but one are already at the metaphase plate.

Oldest known depiction of cells and their nuclei by Antonie van Leeuwenhoek, 1719

Drawing of a Chironomus salivary gland cell published by Walther Flemming in 1882. The nucleus contains polytene chromosomes.
Protein
Videos
Proteins are large biomolecules and macromolecules that comprise one or more long chains of amino acid residues. Proteins perform a vast array of functions within organisms, including catalysing metabolic reactions, DNA replication, responding to stimuli, providing structure to cells and organisms, and transporting molecules from one location to another. Proteins differ from one another primarily in their sequence of amino acids, which is dictated by the nucleotide sequence of their genes, and which usually results in protein folding into a specific 3D structure that determines its activity.

John Kendrew with model of myoglobin in progress

Proteins in different cellular compartments and structures tagged with green fluorescent protein (here, white)

Constituent amino-acids can be analyzed to predict secondary, tertiary and quaternary protein structure, in this case hemoglobin containing heme units