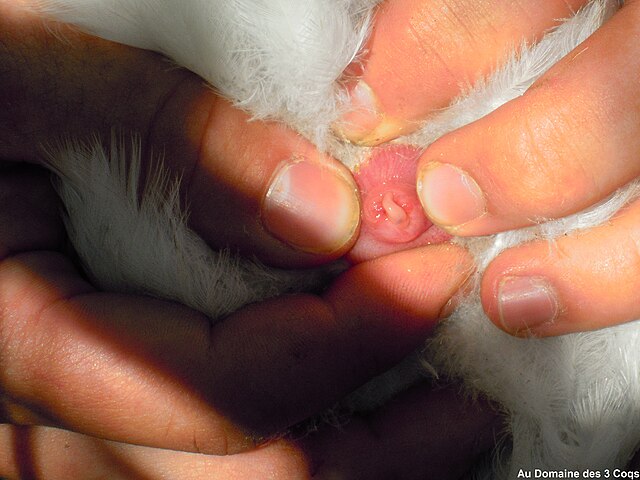
A cloaca, pl.: cloacae, is the rear orifice that serves as the only opening for the digestive, reproductive, and urinary tracts of many vertebrate animals. All amphibians, reptiles, birds, and a few mammals, have this orifice, from which they excrete both urine and feces; this is in contrast to most placental mammals, which have two or three separate orifices for evacuation and reproduction. Excretory openings with analogous purpose in some invertebrates are also sometimes called cloacae. Mating through the cloaca is called cloacal copulation and cloacal kissing.
Cloaca of a red-tailed hawk
Cloaca of a female bird
Cloaca of a male bird
A roseate spoonbill excreting urine in flight
The reproductive system of an organism, also known as the genital system, is the biological system made up of all the anatomical organs involved in sexual reproduction. Many non-living substances such as fluids, hormones, and pheromones are also important accessories to the reproductive system. Unlike most organ systems, the sexes of differentiated species often have significant differences. These differences allow for a combination of genetic material between two individuals, which allows for the possibility of greater genetic fitness of the offspring.
A newborn joey suckles from a teat found within its mother's pouch
A male common frog in nuptial colors waiting for more females to come in a mass of spawn