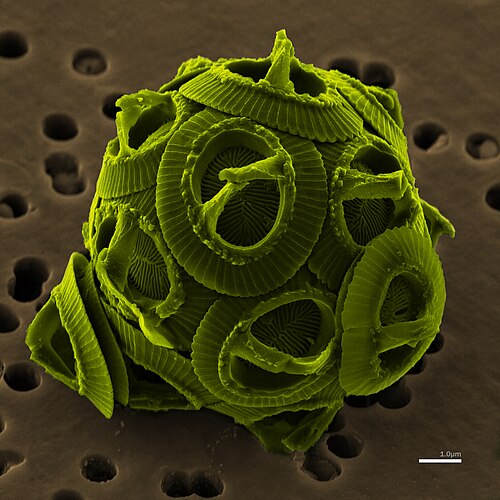
Coccolithophore
Videos
Coccolithophores, or coccolithophorids, are single-celled organisms which are part of the phytoplankton, the autotrophic (self-feeding) component of the plankton community. They form a group of about 200 species, and belong either to the kingdom Protista, according to Robert Whittaker's five-kingdom system, or clade Hacrobia, according to a newer biological classification system. Within the Hacrobia, the coccolithophores are in the phylum or division Haptophyta, class Prymnesiophyceae. Coccolithophores are almost exclusively marine, are photosynthetic, and exist in large numbers throughout the sunlight zone of the ocean.


Satellite photograph: The milky blue colour of this phytoplankton bloom in Barents Sea strongly suggests it contains coccolithophores

A coccolithovirus, Emiliania huxleyi virus 86 (arrowed), infecting an Emiliania huxleyi coccolithophore. This giant marine virus has one of the largest known virus genomes.

Evolutionary history of coccolithophores: (A) Coccolithophore species richness over time; (B) The fossil record of major coccolithophore biomineralization innovations and morphogroups
Hacrobia
Videos
The cryptomonads-haptophytes assemblage is a proposed but disputed monophyletic grouping of unicellular eukaryotes that are not included in the SAR supergroup. Several alternative names have been used for the group, including Hacrobia ; CCTH ; and "Eukaryomonadae".