The Columbian mammoth is an extinct species of mammoth that inhabited the Americas from southern Canada to Costa Rica during the Pleistocene epoch. The Columbian mammoth descended from Eurasian steppe mammoths that colonised North America during the Early Pleistocene around 1.5–1.3 million years ago, and later experienced hybridisation with the woolly mammoth lineage. The Columbian mammoth was among the last mammoth species, and the pygmy mammoths evolved from them on the Channel Islands of California. The closest extant relative of the Columbian and other mammoths is the Asian elephant.
Columbian mammoth
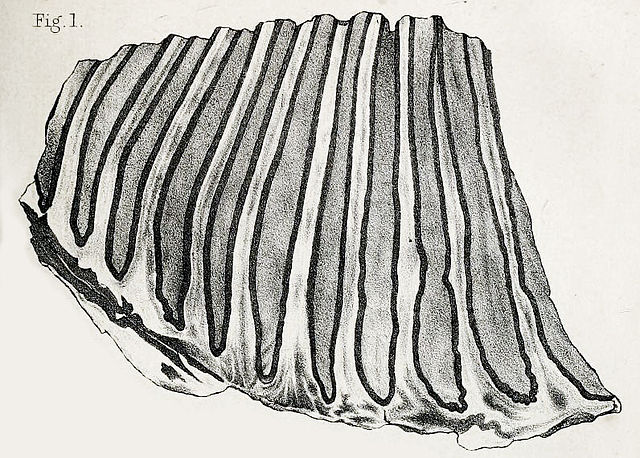
1863 lithograph of the partial holotype molar from Georgia
Henry F. Osborn's 1942 diagram of evolutionarily "progressive" mammoth skulls of species he grouped in the genus Parelephas; the North American species are now considered junior synonyms of M. columbi
Smilodon and dire wolves fighting over a Columbian mammoth carcass in the La Brea Tar Pits, by Robert Bruce Horsfall, 1911
A mammoth is any species of the extinct elephantid genus Mammuthus. They lived from the late Miocene epoch into the Holocene about 4,000 years ago, and various species existed in Africa, Europe, Asia, and North America. Mammoths are distinguished from living elephants by their spirally twisted tusks and in at least some later species, the development of numerous adaptions to living in cold environments, including a thick layer of fur.
Mammoth
Molar of the primitive mammoth Mammuthus meridionalis
Molar of a woolly mammoth
Life restoration of a woolly mammoth at Royal BC Museum