Gorgonopsia is an extinct clade of sabre-toothed therapsids from the Middle to Upper Permian roughly 265 to 252 million years ago. They are characterised by a long and narrow skull, as well as elongated upper and sometimes lower canine teeth and incisors which were likely used as slashing and stabbing weapons. Postcanine teeth are generally reduced or absent. For hunting large prey, they possibly used a bite-and-retreat tactic, ambushing and taking a debilitating bite out of the target, and following it at a safe distance before its injuries exhausted it, whereupon the gorgonopsian would grapple the animal and deliver a killing bite. They would have had an exorbitant gape, possibly in excess of 90°, without having to unhinge the jaw.
Gorgonopsia
Size comparison between a person and Inostrancevia (reconstructed with long lips and some hair)
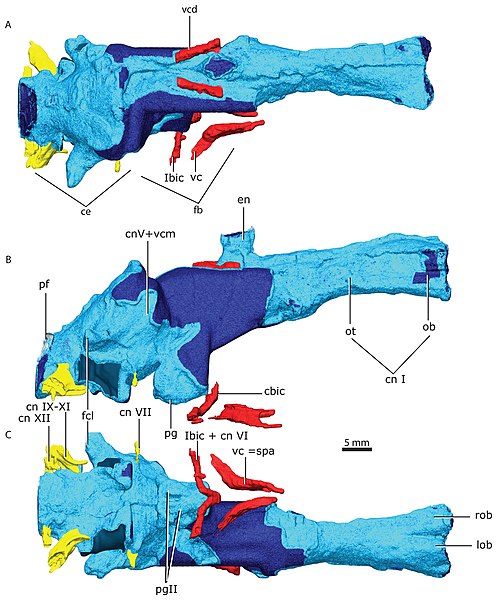
Gorgonopsian brain reconstruction (A. top, B. side, C. underside)
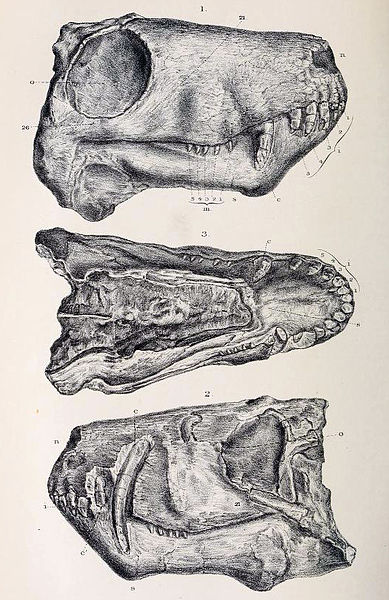
Skull of Aelurosaurus felinus showing tooth arrangement, dual canines, and canine root depth
A saber-tooth is any member of various extinct groups of predatory therapsids, predominantly carnivoran mammals, that are characterized by long, curved saber-shaped canine teeth which protruded from the mouth when closed. Saber-toothed mammals have been found almost worldwide from the Eocene epoch to the end of the Pleistocene epoch.
From top and from left to right, Inostrancevia, Hoplophoneus, Barbourofelis, Smilodon, Machaeroides and Thylacosmilus
1st saber-tooth instance: Gorgonopsidae (Theriodontia, Therapsida, Synapsida) – Lycaenops angusticeps skull
3rd saber-tooth instance: Thylacosmilidae (Sparassodonta) – Thylacosmilus atrox skull
4th saber-tooth instance: Oxyaenidae (Creodonta) – Machaeroides skull