Halteres are a pair of small club-shaped organs on the body of two orders of flying insects that provide information about body rotations during flight. Insects of the large order Diptera (flies) have halteres which evolved from a pair of ancestral hindwings, while males of the much smaller order Strepsiptera (stylops) have halteres which evolved from a pair of ancestral forewings.
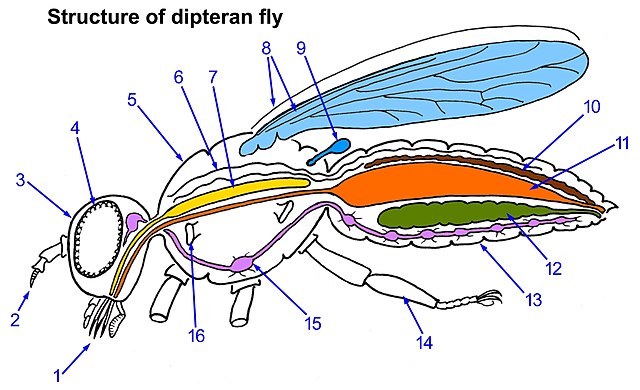
8= single pair of wings and 9= halteres
Crane fly haltere
Electron micrograph of fly haltere and surrounding structures: 1 calyptra (squama) 2 upper calypter (antisquama) 3 haltere 4 mesopleuron 5 hypopleuron 6 coxa 7 wing 8 abdominal segment 9 mesonotum c capitellum of haltere p pedicel of haltere s scabellum of haltere
Haltere length and shape varies from species to species.
Flies are insects of the order Diptera, the name being derived from the Greek δι- di- "two", and πτερόν pteron "wing". Insects of this order use only a single pair of wings to fly, the hindwings having evolved into advanced mechanosensory organs known as halteres, which act as high-speed sensors of rotational movement and allow dipterans to perform advanced aerobatics. Diptera is a large order containing an estimated 1,000,000 species including horse-flies, crane flies, hoverflies, mosquitoes and others, although only about 125,000 species have been described.
Fly
An Anthomyiidae species showing characteristic dipteran features: large eyes, small antennae, sucking mouthparts, single pair of flying wings, hindwings reduced to clublike halteres
Fossil brachyceran in Baltic amber. Lower Eocene, c. 50 million years ago
Fossil nematoceran in Dominican amber. Sandfly, Lutzomyia adiketis (Psychodidae), Early Miocene, c. 20 million years ago