Hypha
Videos
A hypha is a long, branching, filamentous structure of a fungus, oomycete, or actinobacterium. In most fungi, hyphae are the main mode of vegetative growth, and are collectively called a mycelium.

Hyphae of Penicillium

Hyphae growing on tomato sauce
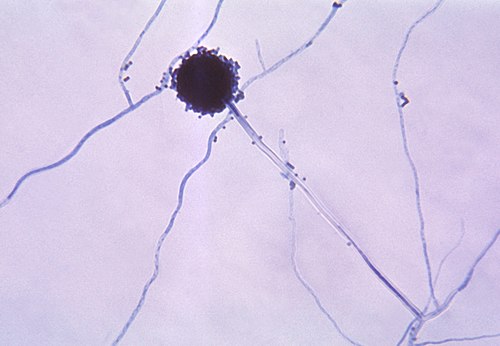
Aspergillus niger
Oomycete
Videos
The Oomycetes, or Oomycota, form a distinct phylogenetic lineage of fungus-like eukaryotic microorganisms within the Stramenopiles. They are filamentous and heterotrophic, and can reproduce both sexually and asexually. Sexual reproduction of an oospore is the result of contact between hyphae of male antheridia and female oogonia; these spores can overwinter and are known as resting spores. Asexual reproduction involves the formation of chlamydospores and sporangia, producing motile zoospores. Oomycetes occupy both saprophytic and pathogenic lifestyles, and include some of the most notorious pathogens of plants, causing devastating diseases such as late blight of potato and sudden oak death. One oomycete, the mycoparasite Pythium oligandrum, is used for biocontrol, attacking plant pathogenic fungi. The oomycetes are also often referred to as water molds, although the water-preferring nature which led to that name is not true of most species, which are terrestrial pathogens.
![Sexual structures (only oogonia, antheridia not shown) of Saprolegnia.[citation needed]](https://upload.wikimedia.org/wikipedia/commons/thumb/e/ef/Oomy1002L.jpg/500px-Oomy1002L.jpg)
Sexual structures (only oogonia, antheridia not shown) of Saprolegnia.[citation needed]

Pythium sp. (Peronosporales), which causes pythiosis in animals, under microscope.

Symptom of late blight (P. infestans, Peronosporales) on the underside of a potato leaf.

Albugo candida (Albuginales) on shepherd's purse (Capsella bursa-pastoris, Brassicales).