Microbial mat
Videos
A microbial mat is a multi-layered sheet of microorganisms, mainly bacteria and archaea, or bacteria alone. Microbial mats grow at interfaces between different types of material, mostly on submerged or moist surfaces, but a few survive in deserts. A few are found as endosymbionts of animals.

The cyanobacterial algal mat, salty lake on the White Sea seaside

Stromatolites are formed by some microbial mats as the microbes slowly move upwards to avoid being smothered by sediment.

Kinneyia-like structure in the Grimsby Formation (Silurian) exposed in Niagara Gorge, New York

Blister-like microbial mat on ripple-marked surface of a Cambrian tidal flat at Blackberry Hill, Wisconsin
Reef
Videos
A reef is a ridge or shoal of rock, coral, or similar relatively stable material lying beneath the surface of a natural body of water. Many reefs result from natural, abiotic (non-living) processes such as deposition of sand or wave erosion planing down rock outcrops. However, reefs such as the coral reefs of tropical waters are formed by biotic (living) processes, dominated by corals and coralline algae. Artificial reefs, such as shipwrecks and other man-made underwater structures, may occur intentionally or as the result of an accident. These are sometimes designed to increase the physical complexity of featureless sand bottoms to attract a more diverse range of organisms. Reefs are often quite near to the surface, but not all definitions require this.

Coral reef at Nusa Lembongan, Bali, Indonesia

Pamalican island with surrounding reef, Sulu Sea, Philippines

A reef surrounding an islet
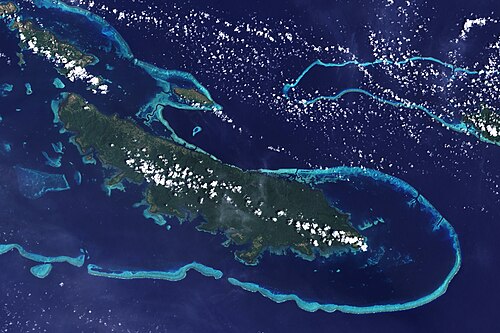
Reefs off Vanatinai Island in the Louisiade Archipelago