Microsporidia are a group of spore-forming unicellular parasites. These spores contain an extrusion apparatus that has a coiled polar tube ending in an anchoring disc at the apical part of the spore. They were once considered protozoans or protists, but are now known to be fungi, or a sister group to fungi. These fungal microbes are obligate eukaryotic parasites that use a unique mechanism to infect host cells. They have recently been discovered in a 2017 Cornell study to infect Coleoptera on a large scale. So far, about 1500 of the probably more than one million species are named. Microsporidia are restricted to animal hosts, and all major groups of animals host microsporidia. Most infect insects, but they are also responsible for common diseases of crustaceans and fish. The named species of microsporidia usually infect one host species or a group of closely related taxa. Approximately 10 percent of the species are parasites of vertebrates —several species, most of which are opportunistic, can infect humans, in whom they can cause microsporidiosis.
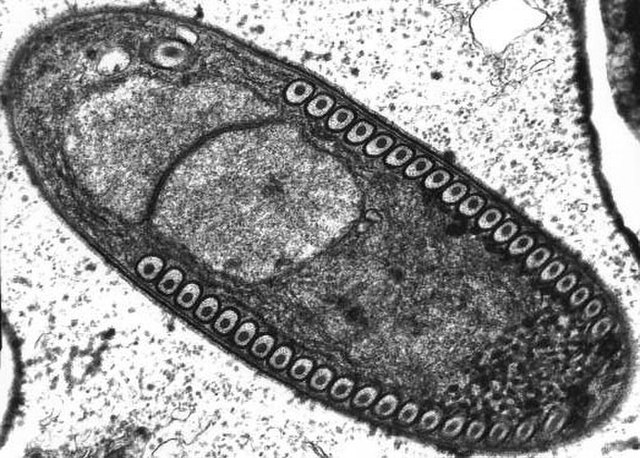
Microsporidia
Xenoma on flatfish caused by Glugea stephani
Dictyocoela diporeiae. A, meront and spore; B, spore wall; C, polar filament
A hyperparasitic microsporidian, Nosema podocotyloidis, a parasite of a digenean which is itself a parasite of a fish.
A unicellular organism, also known as a single-celled organism, is an organism that consists of a single cell, unlike a multicellular organism that consists of multiple cells. Organisms fall into two general categories: prokaryotic organisms and eukaryotic organisms. Most prokaryotes are unicellular and are classified into bacteria and archaea. Many eukaryotes are multicellular, but some are unicellular such as protozoa, unicellular algae, and unicellular fungi. Unicellular organisms are thought to be the oldest form of life, with early protocells possibly emerging 3.8–4.8 billion years ago.
Valonia ventricosa, a species of alga with a diameter that ranges typically from 1 to 4 centimetres (0.4 to 1.6 in) is among the largest unicellular species
Modern stromatolites in Shark Bay, Western Australia. It can take a century for a stromatolite to grow 5 cm.
A bottom-dwelling community found deep in the European Arctic.
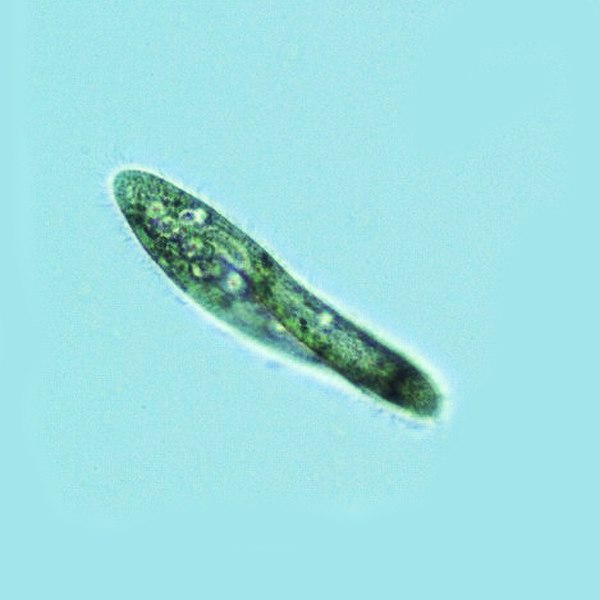
Paramecium tetraurelia, a ciliate, with oral groove visible