Nervous system
Videos
Photos
In biology, the nervous system is the highly complex part of an animal that coordinates its actions and sensory information by transmitting signals to and from different parts of its body. The nervous system detects environmental changes that impact the body, then works in tandem with the endocrine system to respond to such events. Nervous tissue first arose in wormlike organisms about 550 to 600 million years ago. In vertebrates, it consists of two main parts, the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain and spinal cord. The PNS consists mainly of nerves, which are enclosed bundles of the long fibers, or axons, that connect the CNS to every other part of the body. Nerves that transmit signals from the brain are called motor nerves (efferent), while those nerves that transmit information from the body to the CNS are called sensory nerves (afferent). The PNS is divided into two separate subsystems, the somatic and autonomic, nervous systems. The autonomic nervous system is further subdivided into the sympathetic, parasympathetic and enteric nervous systems. The sympathetic nervous system is activated in cases of emergencies to mobilize energy, while the parasympathetic nervous system is activated when organisms are in a relaxed state. The enteric nervous system functions to control the gastrointestinal system. Nerves that exit from the brain are called cranial nerves while those exiting from the spinal cord are called spinal nerves.
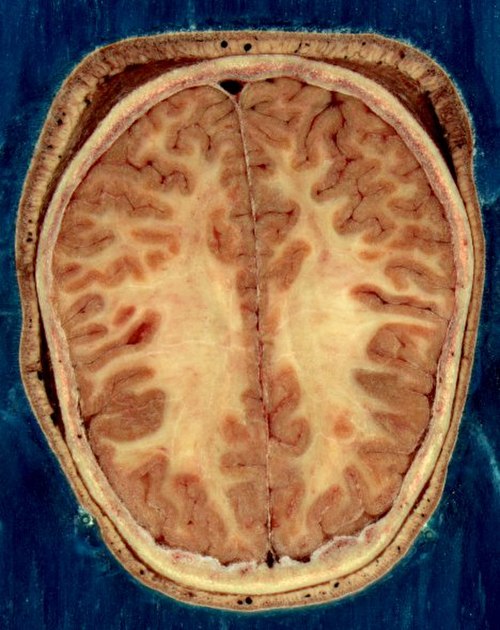
Horizontal section of the head of an adult female human, showing skin, skull, and brain with gray matter (brown in this image) and underlying white matter
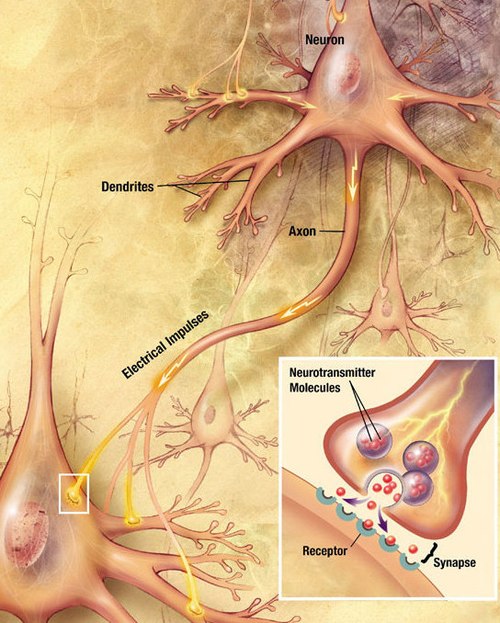
Major elements in synaptic transmission. An electrochemical wave called an action potential travels along the axon of a neuron. When the wave reaches a synapse, it provokes release of a small amount of neurotransmitter molecules, which bind to chemical receptor molecules in the membrane of the target cell.

Illustration of pain pathway, from René Descartes's Treatise of Man

Layers protecting the brain and spinal cord
Sense
Videos
Photos
A sense is a biological system used by an organism for sensation, the process of gathering information about the surroundings through the detection of stimuli. Although, in some cultures, five human senses were traditionally identified as such, many more are now recognized. Senses used by non-human organisms are even greater in variety and number. During sensation, sense organs collect various stimuli for transduction, meaning transformation into a form that can be understood by the brain. Sensation and perception are fundamental to nearly every aspect of an organism's cognition, behavior and thought.

Philippe Mercier - The Sense of Taste - Google Art Project

The sense of smell. Bequest of Mrs E.G. Elgar, 1945 Museum of New Zealand Te Papa Tongarewa.

Lairesse's Allegory of the Five Senses

Detail of The Senses of Hearing, Touch and Taste, Jan Brueghel the Elder, 1618