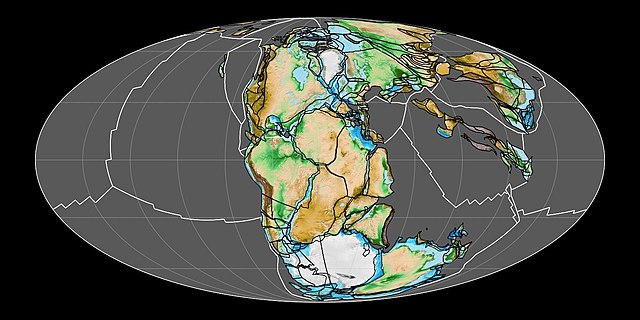
Panthalassa, also known as the Panthalassic Ocean or Panthalassan Ocean, was the vast superocean that encompassed planet Earth and surrounded the supercontinent Pangaea, the latest in a series of supercontinents in the history of Earth. During the Paleozoic–Mesozoic transition, the ocean occupied almost 70% of Earth's surface, with the supercontinent Pangaea taking up less than half. The original, ancient ocean floor has now completely disappeared because of the continuous subduction along the continental margins on its circumference. Panthalassa is also referred to as the Paleo-Pacific or Proto-Pacific because the Pacific Ocean is a direct continuation of Panthalassa.
The Panthalassa superocean 250 million years ago
The supercontinent Pangaea in the early Mesozoic (at 200 Ma) surrounded by Panthalassa.
The Pacific Plate began forming when the triple junction at the center of Panthalassa destabilized about 190 million years ago.
In geology, a supercontinent is the assembly of most or all of Earth's continental blocks or cratons to form a single large landmass. However, some geologists use a different definition, "a grouping of formerly dispersed continents", which leaves room for interpretation and is easier to apply to Precambrian times. To separate supercontinents from other groupings, a limit has been proposed in which a continent must include at least about 75% of the continental crust then in existence in order to qualify as a supercontinent.
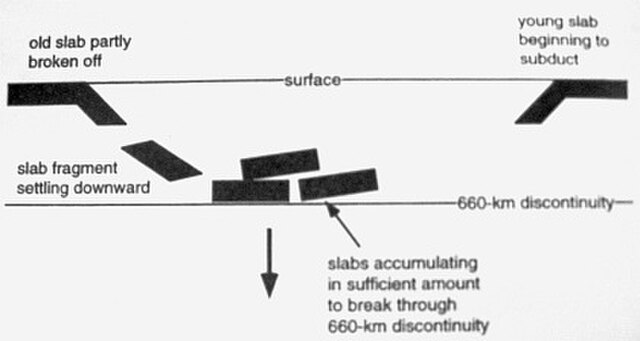
As the slab is subducted into the mantle, the more dense material will break off and sink to the lower mantle creating a discontinuity elsewhere known as a slab avalanche
The effects of mantle plumes possibly caused by slab avalanches elsewhere in the lower mantle on the breakup and assembly of supercontinents
U–Pb ages of 5,246 concordant detrital zircons from 40 of Earth's major rivers