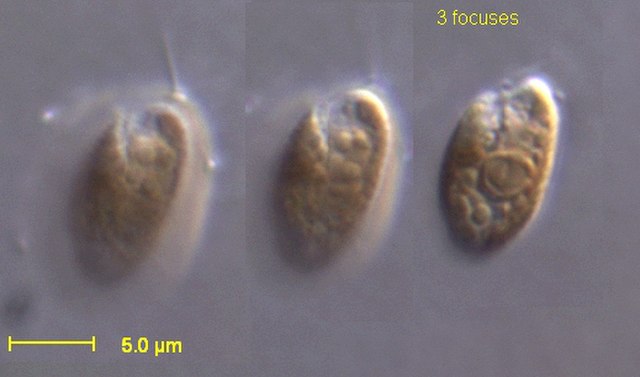
Paramecium is a genus of eukaryotic, unicellular ciliates, commonly studied as a model organism of the ciliate group. Paramecium are widespread in freshwater, brackish, and marine environments and are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. The usefulness of Paramecium as a model organism has caused one ciliate researcher to characterize it as the "white rat" of the phylum Ciliophora.
Paramecium
Paramecia, illustrated by Otto Müller, 1773
Earliest known illustration of Paramecium
"Slipper animalcule," illustrated by Louis Joblot, 1718
The eukaryotes constitute the domain of Eukarya or Eukaryota, organisms whose cells have a membrane-bound nucleus. All animals, plants, fungi, and many unicellular organisms are eukaryotes. They constitute a major group of life forms alongside the two groups of prokaryotes: the Bacteria and the Archaea. Eukaryotes represent a small minority of the number of organisms, but given their generally much larger size, their collective global biomass is much larger than that of prokaryotes.
Image: Rhodomonas salina CCMP 322
Image: Ranunculus asiaticus 4LEST
Image: Trypanosoma sp. PHIL 613 lores
Image: Chaos carolinensis Wilson 1900