Photosynthesis
Videos
Photosynthesis is a system of biological processes by which photosynthetic organisms, such as most plants, algae, and cyanobacteria, convert light energy, typically from sunlight, into the chemical energy necessary to fuel their activities. Photosynthetic organisms use intracellular organic compounds to store the chemical energy they produce in photosynthesis within organic compounds like sugars, glycogen, cellulose and starches. Photosynthesis is usually used to refer to oxygenic photosynthesis, a process that produces oxygen. To use this stored chemical energy, the organisms' cells metabolize the organic compounds through another process called cellular respiration. Photosynthesis plays a critical role in producing and maintaining the oxygen content of the Earth's atmosphere, and it supplies most of the biological energy necessary for complex life on Earth.

Composite image showing the global distribution of photosynthesis, including both oceanic phytoplankton and terrestrial vegetation. Dark red and blue-green indicate regions of high photosynthetic activity in the ocean and on land, respectively.

Plant cells with visible chloroplasts (from a moss, Plagiomnium affine)

Portrait of Jan Baptist van Helmont by Mary Beale, c. 1674

Melvin Calvin works in his photosynthesis laboratory.
Algae
Videos
Algae are any of a large and diverse group of photosynthetic, eukaryotic organisms. The name is an informal term for a polyphyletic grouping that includes species from multiple distinct clades. Included organisms range from unicellular microalgae, such as Chlorella, Prototheca and the diatoms, to multicellular forms, such as the giant kelp, a large brown alga which may grow up to 50 metres (160 ft) in length. Most are aquatic and lack many of the distinct cell and tissue types, such as stomata, xylem and phloem that are found in land plants. The largest and most complex marine algae are called seaweeds, while the most complex freshwater forms are the Charophyta, a division of green algae which includes, for example, Spirogyra and stoneworts. Algae that are carried by water are plankton, specifically phytoplankton.

Image: NSW seabed 1

Image: Водоросли пресноводного водоема 2
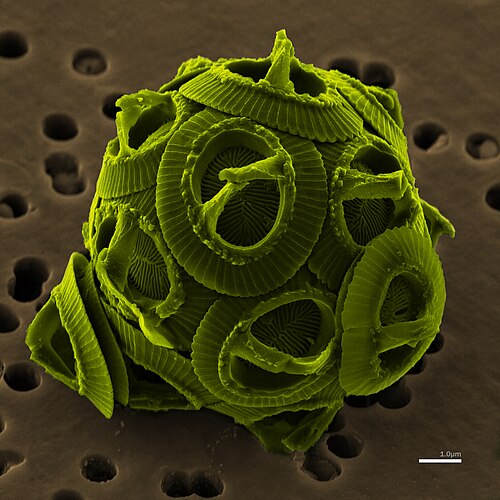
False-color scanning electron micrograph of the unicellular coccolithophore Gephyrocapsa oceanica

The kelp forest exhibit at the Monterey Bay Aquarium: A three-dimensional, multicellular thallus