A plastid, pl. plastids, is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms. They are considered to be intracellular endosymbiotic cyanobacteria.
Plastid
Leucoplasts in plant cells.
An illustration of the stages of inter-conversion in plastids
In cell biology, an organelle is a specialized subunit, usually within a cell, that has a specific function. The name organelle comes from the idea that these structures are parts of cells, as organs are to the body, hence organelle, the suffix -elle being a diminutive. Organelles are either separately enclosed within their own lipid bilayers or are spatially distinct functional units without a surrounding lipid bilayer. Although most organelles are functional units within cells, some function units that extend outside of cells are often termed organelles, such as cilia, the flagellum and archaellum, and the trichocyst.
(A) Electron micrograph of Halothiobacillus neapolitanus cells, arrows highlight carboxysomes. (B) Image of intact carboxysomes isolated from H. neapolitanus. Scale bars are 100 nm.
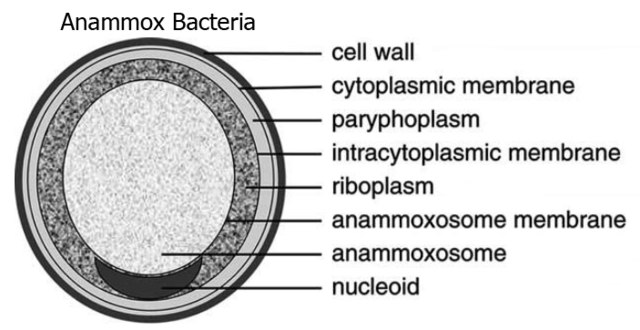
Structure of Candidatus Brocadia anammoxidans, showing an anammoxosome and intracytoplasmic membrane