Seedling
Videos
Page
A seedling is a young sporophyte developing out of a plant embryo from a seed. Seedling development starts with germination of the seed. A typical young seedling consists of three main parts: the radicle, the hypocotyl, and the cotyledons. The two classes of flowering plants (angiosperms) are distinguished by their numbers of seed leaves: monocotyledons (monocots) have one blade-shaped cotyledon, whereas dicotyledons (dicots) possess two round cotyledons. Gymnosperms are more varied. For example, pine seedlings have up to eight cotyledons. The seedlings of some flowering plants have no cotyledons at all. These are said to be acotyledons.

Monocot (left) and dicot (right)

Seedling of a Scots pine

Seedling of Nandina domestica (a dicot) showing two green cotyledon leaves, and the first "true" leaf with its distinct leaflets and red-green color.

A few-days-old Scots pine seedling, the seed still protecting the cotyledons
Embryo
Videos
Page
An embryo is the initial stage of development for a multicellular organism. In organisms that reproduce sexually, embryonic development is the part of the life cycle that begins just after fertilization of the female egg cell by the male sperm cell. The resulting fusion of these two cells produces a single-celled zygote that undergoes many cell divisions that produce cells known as blastomeres. The blastomeres are arranged as a solid ball that when reaching a certain size, called a morula, takes in fluid to create a cavity called a blastocoel. The structure is then termed a blastula, or a blastocyst in mammals.
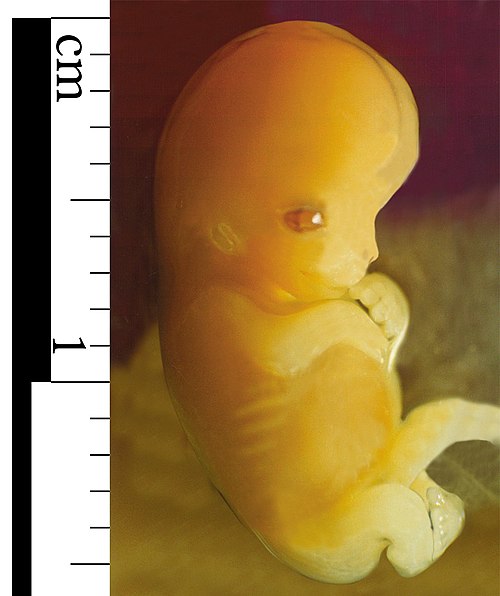
A male human embryo seven weeks after conception (nine weeks gestational age)

Embryos (and one tadpole) of the wrinkled frog (Rana rugosa)

Mouse and snake embryos

The inside of a Ginkgo seed, showing the embryo