Strongyloides stercoralis
Strongyloides stercoralis is a human pathogenic parasitic roundworm causing the disease strongyloidiasis. Its common name in the US is threadworm. In the UK and Australia, however, the term threadworm can also refer to nematodes of the genus Enterobius, otherwise known as pinworms.
Strongyloides stercoralis
Parasitism is a close relationship between species, where one organism, the parasite, lives on or inside another organism, the host, causing it some harm, and is adapted structurally to this way of life. The entomologist E. O. Wilson characterised parasites as "predators that eat prey in units of less than one". Parasites include single-celled protozoans such as the agents of malaria, sleeping sickness, and amoebic dysentery; animals such as hookworms, lice, mosquitoes, and vampire bats; fungi such as honey fungus and the agents of ringworm; and plants such as mistletoe, dodder, and the broomrapes.
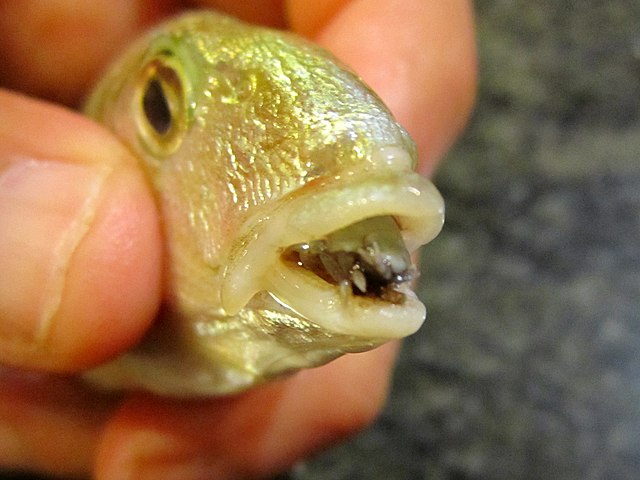
A fish parasite, the isopod Cymothoa exigua, replacing the tongue of a Lithognathus
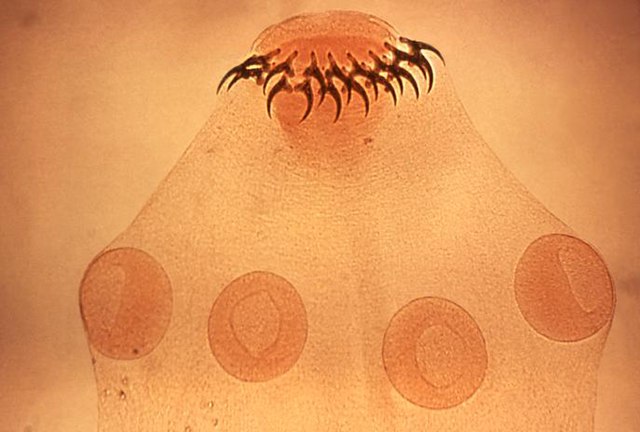
Head (scolex) of tapeworm Taenia solium, an intestinal parasite, has hooks and suckers to attach to its host
The parasitic castrator Sacculina carcini (highlighted) attached to its crab host
Human head-lice are directly transmitted obligate ectoparasites