Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.
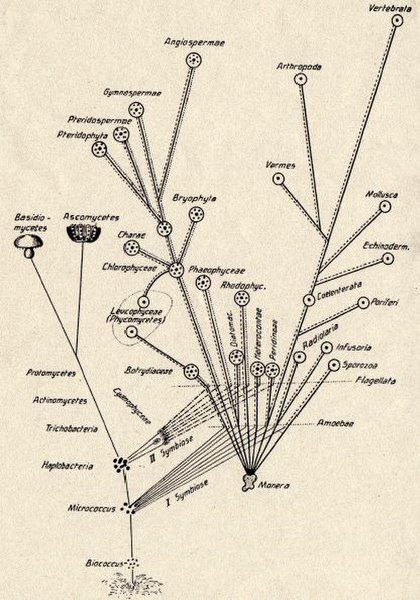
Konstantin Mereschkowski's 1905 tree-of-life diagram, showing the origin of complex life-forms by two episodes of symbiogenesis, the incorporation of symbiotic bacteria to form successively nuclei and chloroplasts
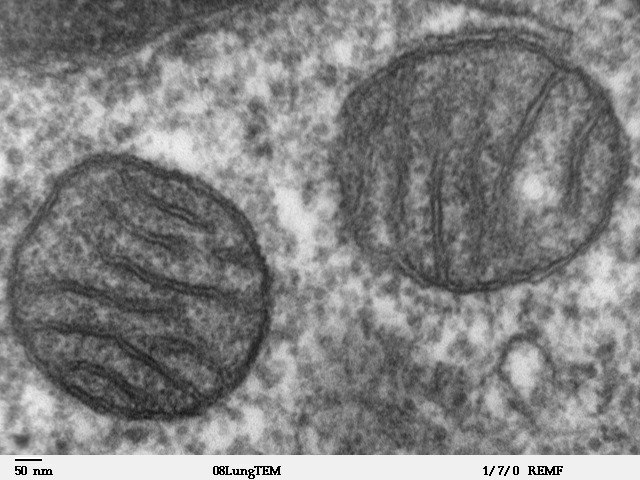
Internal symbiont: mitochondrion has a matrix and membranes, like a free-living alphaproteobacterial cell, from which it may derive.
Evolution is the change in the heritable characteristics of biological populations over successive generations. It occurs when evolutionary processes such as natural selection and genetic drift act on genetic variation, resulting in certain characteristics becoming more or less common within a population over successive generations. The process of evolution has given rise to biodiversity at every level of biological organisation.
White peppered moth
Black morph in peppered moth evolution
Male moor frogs become blue during the height of mating season. Blue reflectance may be a form of intersexual communication. It is hypothesised that males with brighter blue coloration may signal greater sexual and genetic fitness.
The common garter snake has evolved resistance to the defensive substance tetrodotoxin in its amphibian prey.