Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

Lotka–Volterra model of cheetah–baboon interactions. Starting with 80 baboons (green) and 40 cheetahs, this graph shows how the model predicts the two species numbers will progress over time.
Flocks of birds can abruptly change their direction in unison, and then, just as suddenly, make a unanimous group decision to land.
An ecosystem is a system that environments and their organisms form through their interaction. The biotic and abiotic components are linked together through nutrient cycles and energy flows.
Flora of Baja California desert, Cataviña region, Mexico
Global oceanic and terrestrial phototroph abundance, from September 1997 to August 2000. As an estimate of autotroph biomass, it is only a rough indicator of primary production potential and not an actual estimate of it.
A freshwater lake in Gran Canaria, an island of the Canary Islands. Clear boundaries make lakes convenient to study using an ecosystem approach.
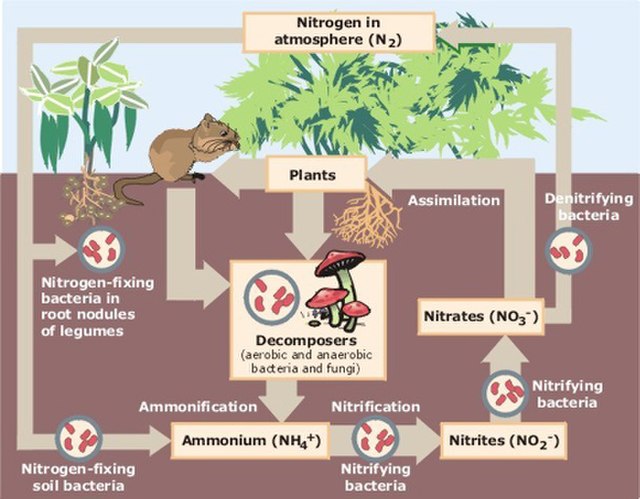
Biological nitrogen cycling