Turtle shell
Videos
Photos
The turtle shell is a shield for the ventral and dorsal parts of turtles, completely enclosing all the vital organs of the turtle and in some cases even the head. It is constructed of modified bony elements such as the ribs, parts of the pelvis and other bones found in most reptiles. The bone of the shell consists of both skeletal and dermal bone, showing that the complete enclosure of the shell likely evolved by including dermal armor into the rib cage.

Scutes (left) and skeletal components (right) of a turtle's carapace. Scutes between the costal (= pleural) and marginals are called supramarginal.
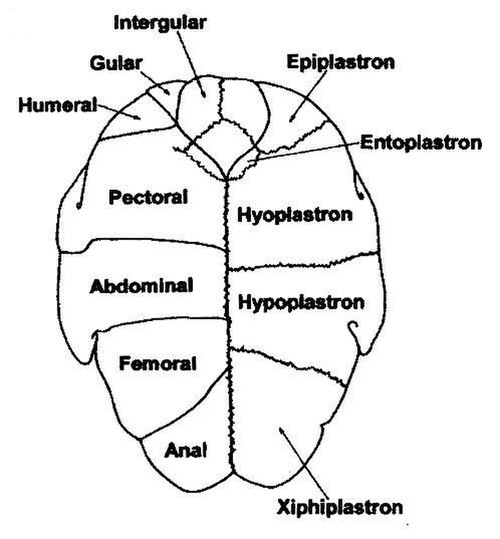
Scutes (left) and skeletal components (right) of a turtle's plastron Pleurodires have an extra scute known as the intergular. It is mostly absent in cryptodires.

Internal anterior carapace of Elseya dentata. Pe=Peripheral, P1=Pleural 1, BCS=Bridge Carapace Suture

Exploded view of the carapace of Emys orbicularis. Legend (i) Neural 1, (ii) Neural 2, (iii) Neural 3, (iv) Neural 4, (v) Neural 5, (vi) Neural 6, (vii) Neural 7, (viii) Neural 8, (ix) extra neural, divided, (x) suprapygal, (xi) nuchal, (xii) right peripheral 1, (xiii) right peripheral 2, (xiv) right peripheral 3, (xv) right peripheral 4, (xvi) right peripheral 5, (xvii) right peripheral 6, (xviii) right peripheral 7, (xix) right peripheral 8, (xx) right peripheral 9, (xxi) right peripheral 10, (xxii) right peripheral 11, (xxiii) pygal, (xxiv) left peripheral 11, (xxv) left peripheral 10, (xxvi) left peripheral 9, (xxvii) left peripheral 8, (xxviii) left peripheral 7, (xxix) left peripheral 6, xxx left peripheral 5, xxxi left peripheral 4, (xxxii) left peripheral 3, (xxxiii) left peripheral 2, (xxxiv) left peripheral 1, (xxxv) right 1st rib, (xxxvi) right pleural 1, (xxxvii) right pleural 2, (xxxviii) right pleural 3, (xxxix) right pleural 4, (xl) right pleural 5, (xli) right pleural 6, (xlii) right pleural 7, (xliii) right pleural 8, (xliv) right 10th rib, (xlv) left 1st rib, (xlvi) left pleural 1, (xlvii) left pleural 2, (xlviii) left pleural 3, (xlix) left pleural 4, (l) left pleural 5, (li) left pleural 6, (lii) left pleural 7, (liii) left pleural 8, (liv) left 10th rib, (9-18) centrums.
Turtle
Videos
Photos
Turtles, or testudines, are reptiles of the order Testudines, characterized by a special shell developed mainly from their ribs. Modern turtles are divided into two major groups, the Pleurodira and Cryptodira, which differ in the way the head retracts. There are 360 living and recently extinct species of turtles, including land-dwelling tortoises and freshwater terrapins. They are found on most continents, some islands and, in the case of sea turtles, much of the ocean. Like other amniotes they breathe air and do not lay eggs underwater, although many species live in or around water.

Turtle

Head and neck of a European pond turtle

Sea turtles have streamlined shells and limbs adapted for fast and efficient swimming.

The red-eared slider has an exceptional seven types of color-detecting cells in its eyes.