Working memory is a cognitive system with a limited capacity that can hold information temporarily. It is important for reasoning and the guidance of decision-making and behavior. Working memory is often used synonymously with short-term memory, but some theorists consider the two forms of memory distinct, assuming that working memory allows for the manipulation of stored information, whereas short-term memory only refers to the short-term storage of information. Working memory is a theoretical concept central to cognitive psychology, neuropsychology, and neuroscience.
Image: Working Memory Label Free
Image: Working Memory Label Free
Memory is the faculty of the mind by which data or information is encoded, stored, and retrieved when needed. It is the retention of information over time for the purpose of influencing future action. If past events could not be remembered, it would be impossible for language, relationships, or personal identity to develop. Memory loss is usually described as forgetfulness or amnesia.
Olin Levi Warner's 1896 illustration, Memory, now housed in the Thomas Jefferson Building at the Library of Congress in Washington, D.C.
The garden of oblivion, illustration by Ephraim Moses Lilien
Regulatory sequence in a promoter at a transcription start site with a paused RNA polymerase and a TOP2B-induced double-strand break
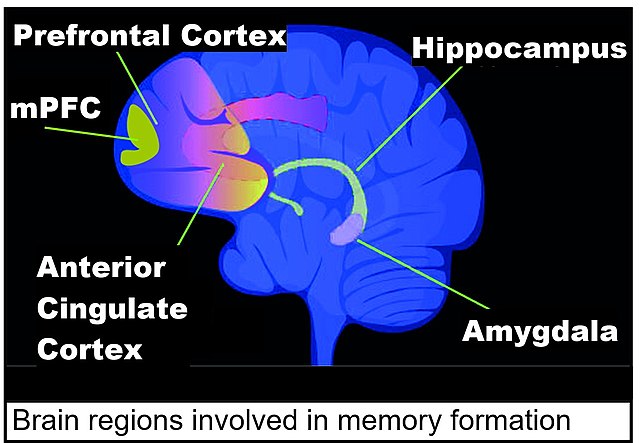
Brain regions involved in memory formation including medial prefrontal cortex (mPFC)