B cell
Videos
Photos
B cells, also known as B lymphocytes, are a type of white blood cell of the lymphocyte subtype. They function in the humoral immunity component of the adaptive immune system. B cells produce antibody molecules which may be either secreted or inserted into the plasma membrane where they serve as a part of B-cell receptors. When a naïve or memory B cell is activated by an antigen, it proliferates and differentiates into an antibody-secreting effector cell, known as a plasmablast or plasma cell. In addition, B cells present antigens and secrete cytokines. In mammals, including marsupials B cells mature in the bone marrow, which is at the core of most bones. In birds, B cells mature in the bursa of Fabricius, a lymphoid organ where they were first discovered by Chang and Glick, which is why the B stands for bursa and not bone marrow, as commonly believed.

Early B cell development: from stem cell to immature B cell
White blood cell
Videos
Photos
White blood cells, also called immune cells or immunocytes, are cells of the immune system that are involved in protecting the body against both infectious disease and foreign invaders. White blood cells include three main subtypes: granulocytes, lymphocytes and monocytes.
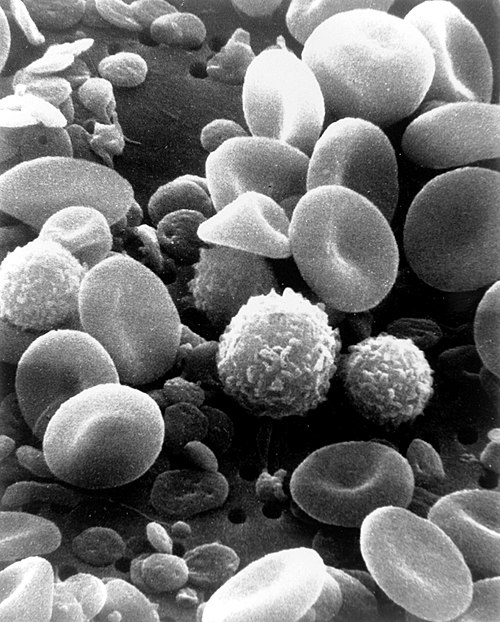
A scanning electron microscope image of normal circulating human blood. In addition to the irregularly shaped leukocytes, both red blood cells and many small disc-shaped platelets are visible.

Neutrophil engulfing anthrax bacteria