Bacteriophage
Videos
Photos
A bacteriophage, also known informally as a phage, is a virus that infects and replicates within bacteria and archaea. The term was derived from "bacteria" and the Greek φαγεῖν, meaning "to devour". Bacteriophages are composed of proteins that encapsulate a DNA or RNA genome, and may have structures that are either simple or elaborate. Their genomes may encode as few as four genes and as many as hundreds of genes. Phages replicate within the bacterium following the injection of their genome into its cytoplasm.

Bacteriophage T2, a member of the Myoviridae due to its contractile tail

Félix d'Herelle conducted the first clinical application of a bacteriophage

George Eliava pioneered the use of phages in treating bacterial infections

In this electron micrograph of bacteriophages attached to a bacterial cell, the viruses are the size and shape of coliphage T1
Archaea
Videos
Photos
Archaea is a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotic. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.

Archaea were discovered in volcanic hot springs. Pictured here is Grand Prismatic Spring of Yellowstone National Park.

The ARMAN are a group of archaea discovered in acid mine drainage in the early 2000s.
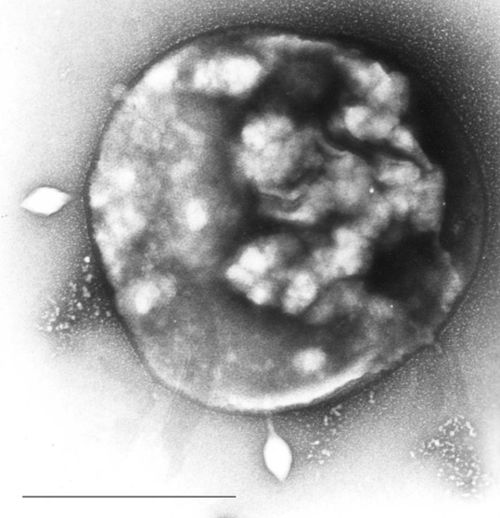
Sulfolobus infected with the DNA virus STSV1. Bar is 1 micrometer.

Archaea that grow in the hot water of the Morning Glory Hot Spring in Yellowstone National Park produce a bright colour