Fructose 1,6-bisphosphatase
Videos
The enzyme fructose bisphosphatase (EC 3.1.3.11; systematic name D-fructose-1,6-bisphosphate 1-phosphohydrolase) catalyses the conversion of fructose-1,6-bisphosphate to fructose 6-phosphate in gluconeogenesis and the Calvin cycle, which are both anabolic pathways:D-fructose 1,6-bisphosphate + H2O = D-fructose 6-phosphate + phosphate

crystal structure of rabbit liver fructose-1,6-bisphosphatase at 2.3 angstrom resolution

crystal structure of fructose-1,6-bisphosphatase

AMP
Glycolysis
Videos
Glycolysis is the metabolic pathway that converts glucose into pyruvate and, in most organisms, occurs in the liquid part of cells. The free energy released in this process is used to form the high-energy molecules adenosine triphosphate (ATP) and reduced nicotinamide adenine dinucleotide (NADH). Glycolysis is a sequence of ten reactions catalyzed by enzymes.
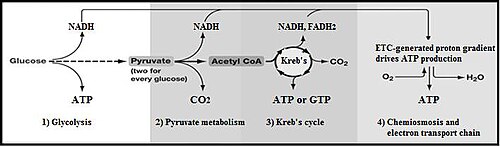
Summary of aerobic respiration

Eduard Buchner. Discovered cell-free fermentation.

Otto Meyerhof. One of the main scientists involved in completing the puzzle of glycolysis