Green fluorescent protein
Videos
The green fluorescent protein (GFP) is a protein that exhibits green fluorescence when exposed to light in the blue to ultraviolet range. The label GFP traditionally refers to the protein first isolated from the jellyfish Aequorea victoria and is sometimes called avGFP. However, GFPs have been found in other organisms including corals, sea anemones, zoanithids, copepods and lancelets.

Structure of the Aequorea victoria green fluorescent protein.

Aequorea victoria
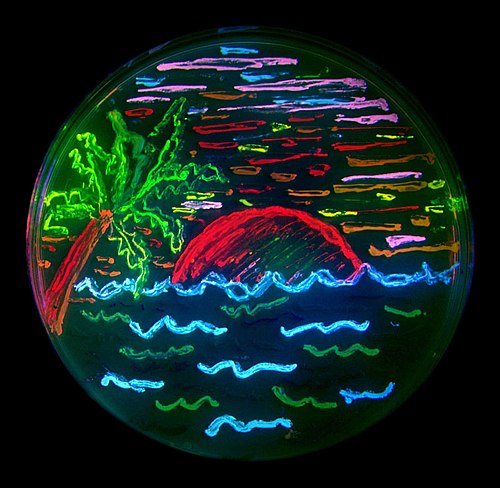
The diversity of genetic mutations is illustrated by this San Diego beach scene drawn with living bacteria expressing 8 different colors of fluorescent proteins (derived from GFP and dsRed).

Different proteins produce different fluorescent colors when exposed to ultraviolet light.
Roger Y. Tsien
Videos
Roger Yonchien Tsien was an American biochemist. He was a professor of chemistry and biochemistry at the University of California, San Diego and was awarded the Nobel Prize in Chemistry in 2008 for his discovery and development of the green fluorescent protein, in collaboration with organic chemist Osamu Shimomura and neurobiologist Martin Chalfie. Tsien was also a pioneer of calcium imaging.


Paul Krugman, Roger Tsien, Martin Chalfie, Osamu Shimomura, Makoto Kobayashi and Toshihide Masukawa, Nobel Prize Laureates 2008, at a press conference at the Swedish Academy of Science in Stockholm.