Grey matter, or brain matter in American English, is a major component of the central nervous system, consisting of neuronal cell bodies, neuropil, glial cells, synapses, and capillaries. Grey matter is distinguished from white matter in that it contains numerous cell bodies and relatively few myelinated axons, while white matter contains relatively few cell bodies and is composed chiefly of long-range myelinated axons. The colour difference arises mainly from the whiteness of myelin. In living tissue, grey matter actually has a very light grey colour with yellowish or pinkish hues, which come from capillary blood vessels and neuronal cell bodies.
Micrograph showing grey matter, with the characteristic neuronal cell bodies (dark shade of pink), and white matter with its characteristic fine meshwork-like appearance (left of image; lighter shade of pink). HPS stain.
Human brain right dissected lateral view
An axon or nerve fiber is a long, slender projection of a nerve cell, or neuron, in vertebrates, that typically conducts electrical impulses known as action potentials away from the nerve cell body. The function of the axon is to transmit information to different neurons, muscles, and glands. In certain sensory neurons, such as those for touch and warmth, the axons are called afferent nerve fibers and the electrical impulse travels along these from the periphery to the cell body and from the cell body to the spinal cord along another branch of the same axon. Axon dysfunction can be the cause of many inherited and acquired neurological disorders that affect both the peripheral and central neurons. Nerve fibers are classed into three types – group A nerve fibers, group B nerve fibers, and group C nerve fibers. Groups A and B are myelinated, and group C are unmyelinated. These groups include both sensory fibers and motor fibers. Another classification groups only the sensory fibers as Type I, Type II, Type III, and Type IV.

A dissected human brain, showing grey matter and white matter
TEM of a myelinated axon in cross-section.
Axon of nine-day-old mouse with growth cone visible
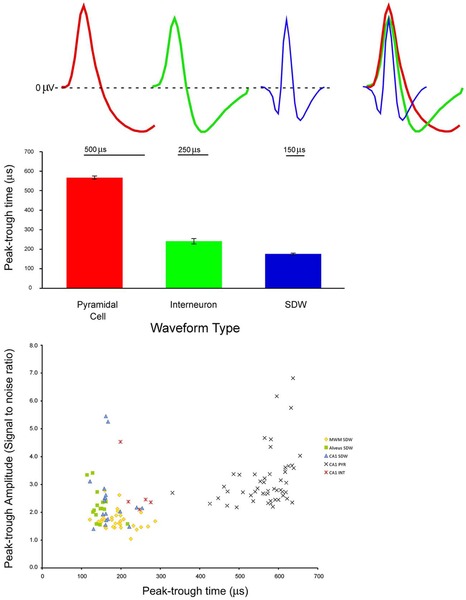
(A) pyramidal cell, interneuron, and short duration waveform (Axon), overlay of the three average waveforms; (B) Average and standard error of peak-trough time for pyramidal cells interneurons, and putative axons; (C) Scatter plot of signal to noise ratios for individual units againstpeak-trough time for axons, pyramidal cells (PYRs) and interneurons (INTs).