Macrophage
Videos
Macrophages are a type of white blood cell of the innate immune system that engulf and digest pathogens, such as cancer cells, microbes, cellular debris, and foreign substances, which do not have proteins that are specific to healthy body cells on their surface. This process is called phagocytosis, which acts to defend the host against infection and injury.

Gram stain of a macrophage with ingested S. epidermidis bacteria, seen as purple granules within its cytoplasm.

A macrophage stretching its "arms" (filopodia) to engulf two particles, possibly pathogens, in a mouse (trypan blue exclusion staining).

Siderophage

Anthracotic macrophage
Adaptive immune system
Videos
The adaptive immune system, also known as the acquired immune system, or specific immune system is a subsystem of the immune system that is composed of specialized, systemic cells and processes that eliminate pathogens or prevent their growth. The acquired immune system is one of the two main immunity strategies found in vertebrates.
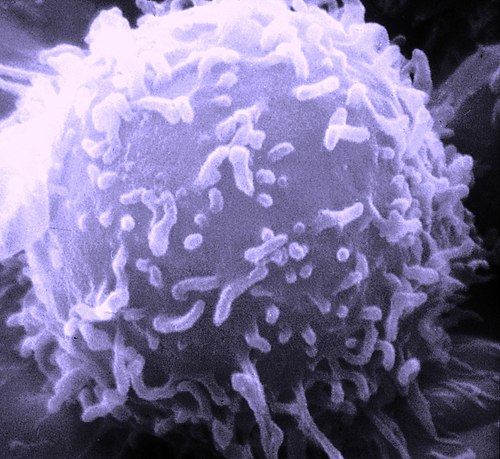
A scanning electron microscope image of a single human lymphocyte