Monito del monte
Videos
Photos
The monito del monte , or colocolo opossum, is a diminutive species of marsupial native only to south-western South America. It is the only extant species in the ancient order Microbiotheria, and the sole New World representative of the superorder Australidelphia, being more closely related to Australian marsupials than to other American marsupials. The species is nocturnal and arboreal, and lives in thickets of South American mountain bamboo in the Valdivian temperate forests of the southern Andes, aided by its partially prehensile tail. It consumes an omnivorous diet based on insects and fruit.

Monito del monte

A monito del monte in state of torpor.
Marsupial
Videos
Photos
Marsupials are a diverse group of mammals belonging to the infraclass Marsupialia. They are primarily found in Australasia, Wallacea, and the Americas. One of the defining features of marsupials is their unique reproductive strategy, where the young are born in a relatively undeveloped state and then nurtured within a pouch.

Marsupial

Koala (Phascolarctos cinereus)

Female eastern grey kangaroo with a joey in her pouch
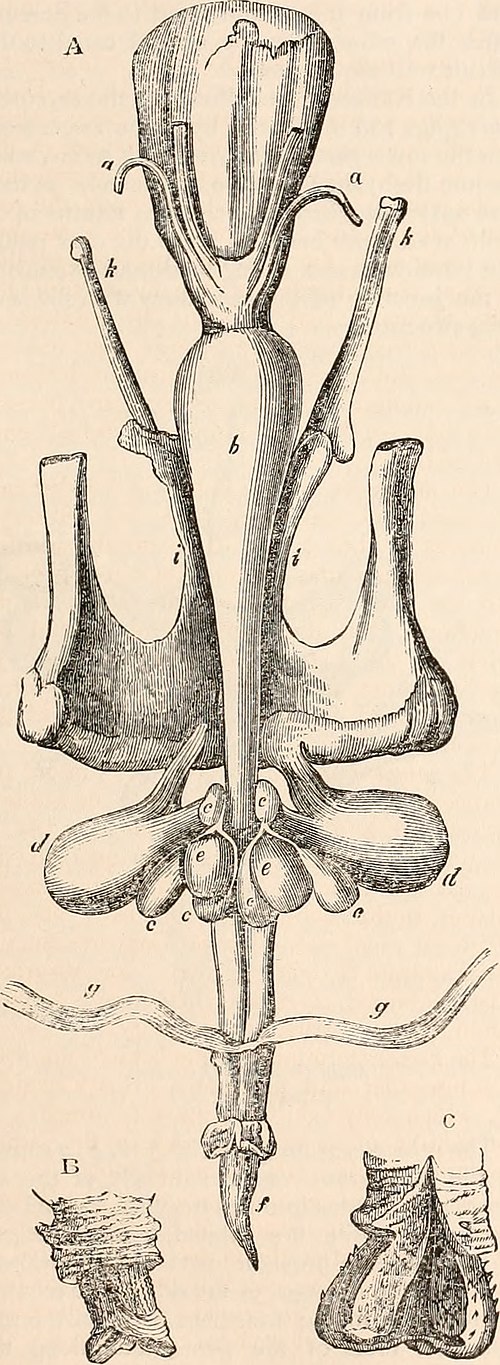
Reproductive tract of a male macropod