Neutrophil
Videos
Neutrophils are a type of white blood cell. More specifically, they form the most abundant type of granulocytes and make up 40% to 70% of all white blood cells in humans. They form an essential part of the innate immune system, with their functions varying in different animals.
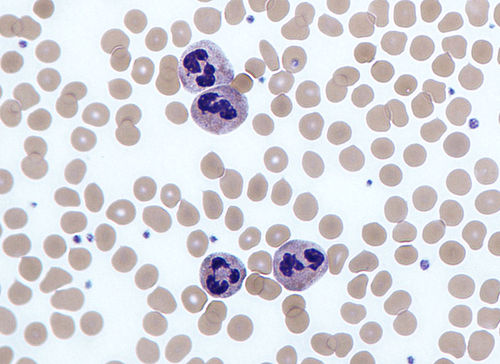
Neutrophils with segmented nuclei surrounded by erythrocytes and platelets. Intra-cellular granules are visible in the cytoplasm (Giemsa stained).

Hypersegmented neutrophil

Scanning electron micrograph of a neutrophil (yellow) phagocytosing anthrax bacilli (orange). Scale bar is 5 μm.

Micrograph showing several neutrophils during an acute inflammation
Granulocyte
Videos
Granulocytes are cells in the innate immune system characterized by the presence of specific granules in their cytoplasm. Such granules distinguish them from the various agranulocytes. All myeloblastic granulocytes are polymorphonuclear, that is, they have varying shapes (morphology) of the nucleus ; and are referred to as polymorphonuclear leukocytes. In common terms, polymorphonuclear granulocyte refers specifically to "neutrophil granulocytes", the most abundant of the granulocytes; the other types have varying morphology. Granulocytes are produced via granulopoiesis in the bone marrow.

Types of granulocytes

A neutrophil with a segmented nucleus (center and surrounded by erythrocytes), the intra-cellular granules are visible in the cytoplasm (Giemsa-stained high magnification)