Reptiliomorpha is a clade containing the amniotes and those tetrapods that share a more recent common ancestor with amniotes than with living amphibians (lissamphibians). It was defined by Michel Laurin (2001) and Vallin and Laurin (2004) as the largest clade that includes Homo sapiens, but not Ascaphus truei. Laurin and Reisz (2020) defined Pan-Amniota as the largest total clade containing Homo sapiens, but not Pipa pipa, Caecilia tentaculata, and Siren lacertina.
Limnoscelis, a carnivorous diadectomorph
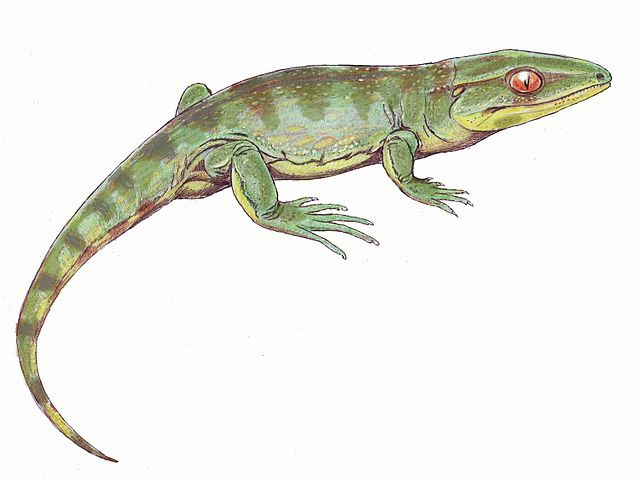
Gephyrostegus, a small terrestrial tetrapod
Discosauriscus, a neotenic seymouriamorph
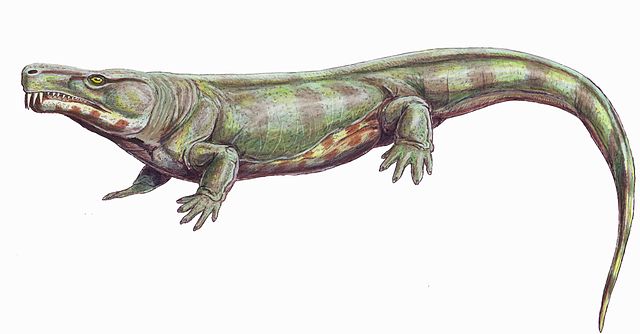
Archeria, an aquatic embolomere
Amniotes are tetrapod vertebrate animals belonging to the clade Amniota, a large group that comprises the vast majority of living terrestrial and semiaquatic vertebrates. Amniotes evolved from amphibian ancestors during the Carboniferous period and further diverged into two groups, namely the sauropsids and synapsids. They are distinguished from the other living tetrapod clade — the non-amniote lissamphibians — by the development of three extraembryonic membranes, thicker and keratinized skin, and costal respiration.
Amniote
Archaeothyris, one of the most basal synapsids, first appears in the fossil records about 306 million years ago.
By the Mesozoic, 150 million years ago, sauropsids included the largest animals anywhere. Shown are some late Jurassic dinosaurs, including the early bird Archaeopteryx perched on a tree stump.