Skeletal muscle
Videos
Skeletal muscles are organs of the vertebrate muscular system and typically are attached by tendons to bones of a skeleton. The muscle cells of skeletal muscles are much longer than in the other types of muscle tissue, and are often known as muscle fibers. The muscle tissue of a skeletal muscle is striated – having a striped appearance due to the arrangement of the sarcomeres.
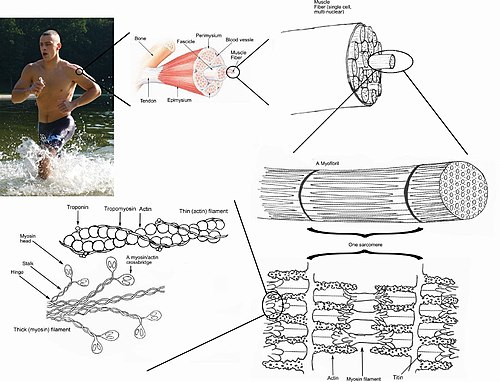
A top-down view of skeletal muscle

ATPase staining of a muscle cross section. Type II fibers are dark, due to the alkaline pH of the preparation. In this example, the size of the type II fibers is considerably less than the type I fibers due to denervation atrophy.

Diagram of sarcoplasmic reticulum with terminal cisternae and T-tubules.

When a sarcomere contracts, the Z lines move closer together, and the I band becomes smaller. The A band stays the same width. At full contraction, the thin and thick filaments overlap.
Tendon
Videos
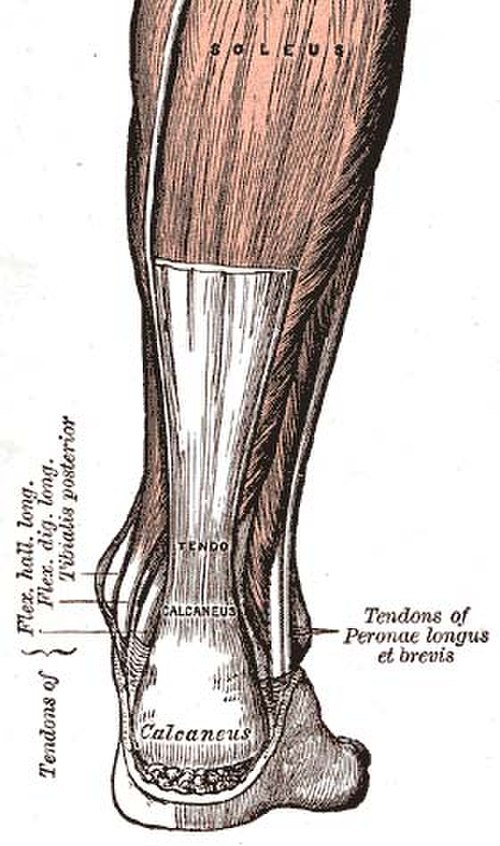
A tendon or sinew is a tough band of dense fibrous connective tissue that connects muscle to bone. It sends the mechanical forces of muscle contraction to the skeletal system, while withstanding tension.
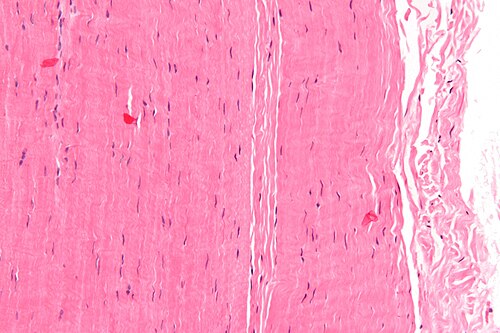
Micrograph of a piece of tendon; H&E stain
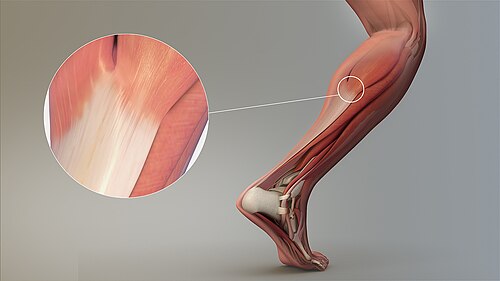
Magnified view of a tendon

Ossified tendon from an Edmontosaurus bone bed in Wyoming (Lance Formation)