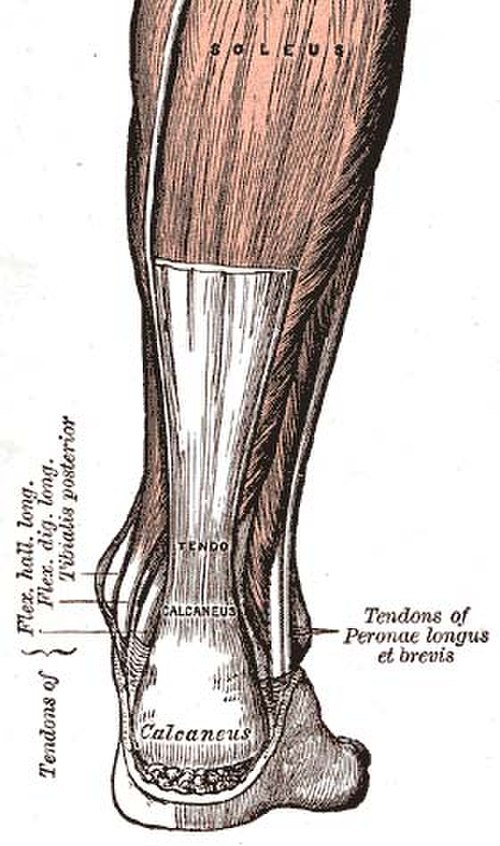
Tendon
Videos
A tendon or sinew is a tough band of dense fibrous connective tissue that connects muscle to bone. It sends the mechanical forces of muscle contraction to the skeletal system, while withstanding tension.
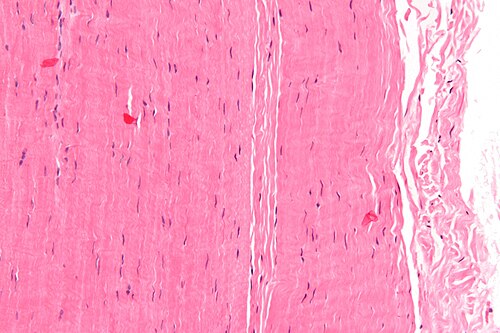
Micrograph of a piece of tendon; H&E stain
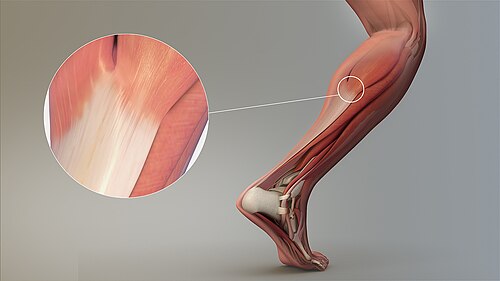
Magnified view of a tendon

Ossified tendon from an Edmontosaurus bone bed in Wyoming (Lance Formation)
Collagen
Videos
Collagen is the main structural protein in the extracellular matrix found in the body's various connective tissues. As the main component of connective tissue, it is the most abundant protein in mammals, making up from 25% to 35% of the whole-body protein content. Collagen consists of amino acids bound together to form a triple helix of elongated fibril known as a collagen helix. It is mostly found in connective tissue such as cartilage, bones, tendons, ligaments, and skin. Vitamin C is vital for collagen synthesis, and Vitamin E improves the production of collagen.


Collagen

The D-period of collagen fibrils results in visible 67nm bands when observed by electron microscopy.

A salami and the collagen casing (below) it came in