Three-spined stickleback
Videos
Page
The three-spined stickleback is a fish native to most inland and coastal waters north of 30°N. It has long been a subject of scientific study for many reasons. It shows great morphological variation throughout its range, ideal for questions about evolution and population genetics. Many populations are anadromous and very tolerant of changes in salinity, a subject of interest to physiologists. It displays elaborate breeding behavior and it can be social making it a popular subject of inquiry in fish ethology and behavioral ecology. Its antipredator adaptations, host-parasite interactions, sensory physiology, reproductive physiology, and endocrinology have also been much studied. Facilitating these studies is the fact that the three-spined stickleback is easy to find in nature and easy to keep in aquaria.

Three-spined stickleback

Male (redish) and a gravid female three-spined stickleback

Male stickleback with red throat and shiny blue eye

Reproduction of the three-spined stickleback
Fish migration
Videos
Page
Fish migration is mass relocation by fish from one area or body of water to another. Many types of fish migrate on a regular basis, on time scales ranging from daily to annually or longer, and over distances ranging from a few metres to thousands of kilometres. Such migrations are usually done for better feeding or to reproduce, but in other cases the reasons are unclear.

Many species of salmon are anadromous and can migrate long distances up rivers to spawn
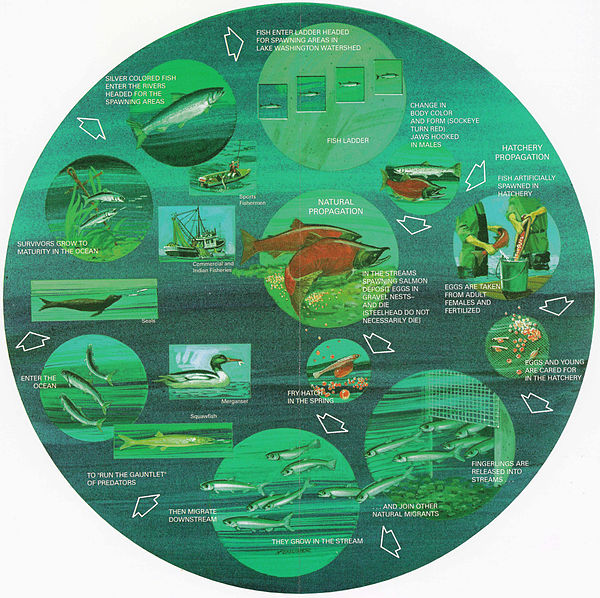
Life cycle of anadromous fish. From a U.S. Government pamphlet. (Click image to enlarge.)